miRZip™ Pooled Anti-miRNA Virus Library

Products
| Catalog Number | Description | Size | Price | Quantity | Add to Cart | |||
|---|---|---|---|---|---|---|---|---|
| MZIPPLVA-1 | miRZip™ Pooled Anti-miRNA Virus Library | 1 Virus Aliquot | $1412 |
|
||||
Overview
Overview
Simplifying high-throughput phenotypic screening for miR function
Take your miR studies to the next level with SBI’s pooled miRZip™ anti-miRNA Lentivirus Library. Quickly and efficiently create stable miR knockdowns in the cell line of your choice for high-throughput phenotypic screening. SBI’s miRZip constructs leverage our powerful and well-regarded third-generation lentivector technology to deliver short hairpin RNAs (shRNA) that, after processing, preferentially produce an anti-sense miRNA. The hairpin is rationally designed to be asymmetric, ensuring that the sense strand does not contain the endogenous miRNA sequence and enabling accumulation of the anti-miRNA. The result is robust derepression of the transcripts targeted by the microRNA being "zipped,” and elevation of the corresponding protein levels.
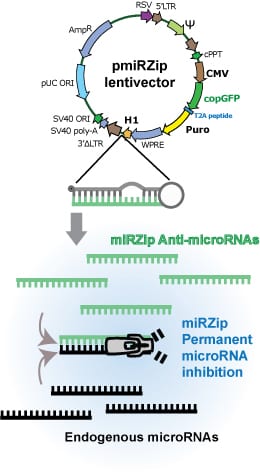 Study miRs with SBI's powerful miRZip technology:
Study miRs with SBI's powerful miRZip technology:
- Stable and permanent anti-microRNA expression from a constitutive H1 promoter
- Rationally designed, asymmetric hairpins optimized for anti-sense microRNA production
- Efficient suppression of specific endogenous microRNAs
- Reliable delivery to dividing or non-dividing cells
- Selection/sorting for transfected/transduced cells with either copGFP or puromycin
References
How It Works
How It Works
Screening with the miRZip Library is straightforward
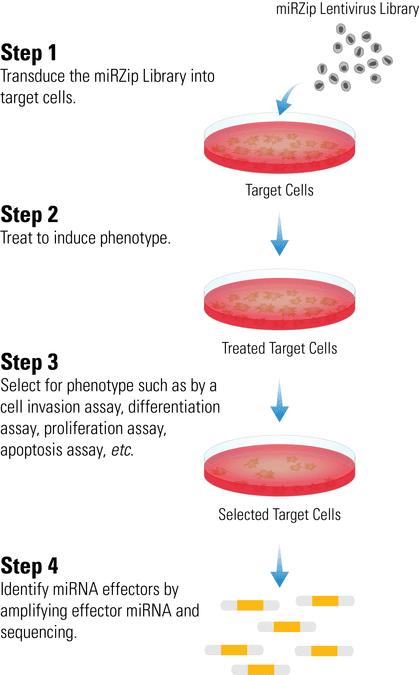
- Transduce the miRZip Library into target cells
- Treat to induce phenotype
- Use a functional assay (invasion assay, differentiation assay, proliferation assay, apoptosis assay, etc.) to select for cells exhibiting the desired phenotype
- Identify miR effectors by amplifying effector miR and sequencing
Supporting Data
Supporting Data
See SBI’s mirZip technology in action
miRZip constructs express high levels of anti-microRNAs
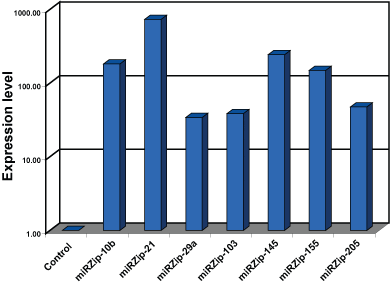
Figure 1. miRZip constructs express high levels of anti-microRNAs. miRZip constructs were transfected into HEK293 cells, total RNA isolated, and miRZip levels measured using SBI’s QuantiMir Assay (Cat.# RA420A-1). Compared to QuantiMir assays performed on RNA isolated from non-transfected cells, the miRZip-transfected cells delivered high expression levels of the specific miRZip anti-miRNAs.
Efficient modulation of target protein levels by SBI’s Lenti-miR and miRZip technologies 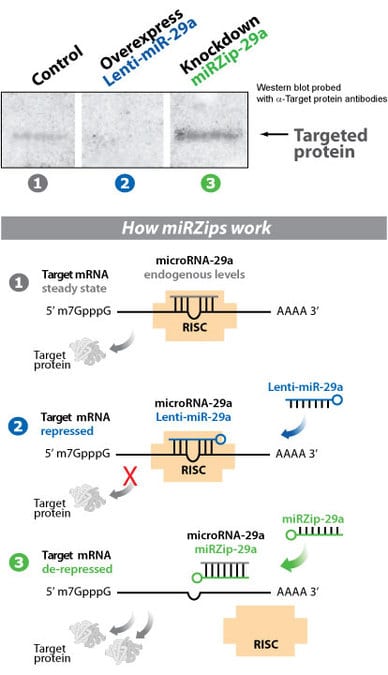
Figure 2. Target protein levels are efficiently modulated by SBI’s Lenti-miR-29a and miRZip-29a microRNA constructs.
miRZip technology is effective in a functional assay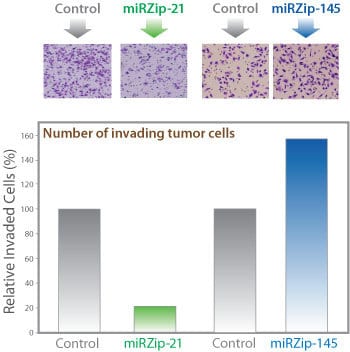
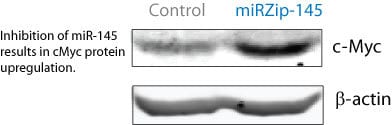
Figure 3. miRZip-21 and miRZip-145 exhibit anti-oncogenic and oncogenic activity, respectively, in a cell invasion assay using MDA-MB-231 breast cancer cells. (Top panel) miR-21 is an oncogenic miRNA and miR-145 is a tumor suppressing miRNA. Transduction of miRZip-21 (the anti-miR21 lentiviral construct) and miRZip-145 (the anti-miR145 lentiviral construct), inhibited the respective miRs, resulting in a lower percentage of invading cells in miRZip-21-transduced cells and a higher percentage of invading cells in miRZip-145-transduced cells. (Bottom panel) Additionally, the miRZip-145 lentivirus inhibited endogenous miR-145 and elevated protein expression levels of the miR-145 target oncogene c-Myc.
FAQs
Documentation
Citations
Related Products
Products
| Catalog Number | Description | Size | Price | Quantity | Add to Cart | |||
|---|---|---|---|---|---|---|---|---|
| MZIPPLVA-1 | miRZip™ Pooled Anti-miRNA Virus Library | 1 Virus Aliquot | $1412 |
|
||||
Overview
Overview
Simplifying high-throughput phenotypic screening for miR function
Take your miR studies to the next level with SBI’s pooled miRZip™ anti-miRNA Lentivirus Library. Quickly and efficiently create stable miR knockdowns in the cell line of your choice for high-throughput phenotypic screening. SBI’s miRZip constructs leverage our powerful and well-regarded third-generation lentivector technology to deliver short hairpin RNAs (shRNA) that, after processing, preferentially produce an anti-sense miRNA. The hairpin is rationally designed to be asymmetric, ensuring that the sense strand does not contain the endogenous miRNA sequence and enabling accumulation of the anti-miRNA. The result is robust derepression of the transcripts targeted by the microRNA being "zipped,” and elevation of the corresponding protein levels.
Study miRs with SBI's powerful miRZip technology:
- Stable and permanent anti-microRNA expression from a constitutive H1 promoter
- Rationally designed, asymmetric hairpins optimized for anti-sense microRNA production
- Efficient suppression of specific endogenous microRNAs
- Reliable delivery to dividing or non-dividing cells
- Selection/sorting for transfected/transduced cells with either copGFP or puromycin
References
How It Works
How It Works
Screening with the miRZip Library is straightforward
- Transduce the miRZip Library into target cells
- Treat to induce phenotype
- Use a functional assay (invasion assay, differentiation assay, proliferation assay, apoptosis assay, etc.) to select for cells exhibiting the desired phenotype
- Identify miR effectors by amplifying effector miR and sequencing
Supporting Data
Supporting Data
See SBI’s mirZip technology in action
miRZip constructs express high levels of anti-microRNAs
Figure 1. miRZip constructs express high levels of anti-microRNAs. miRZip constructs were transfected into HEK293 cells, total RNA isolated, and miRZip levels measured using SBI’s QuantiMir Assay (Cat.# RA420A-1). Compared to QuantiMir assays performed on RNA isolated from non-transfected cells, the miRZip-transfected cells delivered high expression levels of the specific miRZip anti-miRNAs.
Efficient modulation of target protein levels by SBI’s Lenti-miR and miRZip technologies
Figure 2. Target protein levels are efficiently modulated by SBI’s Lenti-miR-29a and miRZip-29a microRNA constructs.
miRZip technology is effective in a functional assay
Figure 3. miRZip-21 and miRZip-145 exhibit anti-oncogenic and oncogenic activity, respectively, in a cell invasion assay using MDA-MB-231 breast cancer cells. (Top panel) miR-21 is an oncogenic miRNA and miR-145 is a tumor suppressing miRNA. Transduction of miRZip-21 (the anti-miR21 lentiviral construct) and miRZip-145 (the anti-miR145 lentiviral construct), inhibited the respective miRs, resulting in a lower percentage of invading cells in miRZip-21-transduced cells and a higher percentage of invading cells in miRZip-145-transduced cells. (Bottom panel) Additionally, the miRZip-145 lentivirus inhibited endogenous miR-145 and elevated protein expression levels of the miR-145 target oncogene c-Myc.