pCDH-CuO-MCS SparQ™ Cloning and Expression Lentivector
- Robust—increase expression up to 32-fold
- Adjustable—tune expression levels by titrating the amount of cumate
- Reversible—turn expression on, then off, then on again
- Versatile—choose from all-in-one formats that co-express CymR and your gene-of-interest, or two-vector systems where CymR is expressed from a different plasmid
- Powerful—suitable for in vivo applications

Products
| Catalog Number | Description | Size | Price | Quantity | Add to Cart | |||
|---|---|---|---|---|---|---|---|---|
| QM500A-1 | pCDH-CuO-MCS empty cloning lentivector | 10 µg | $554 |
|
||||
Overview
Overview
Get tight, titratable gene expression
With SBI’s SparQ™ Cumate Switch System, you can get inducible gene expression in mammalian cells with a range of cloning and expression lentivectors. The pCDH-CuO-MCS SparQ™ Cloning and Expression Lentivector maximizes your flexibility with only an MCS downstream of the cumate switch promoter—clone-in your gene of interest and the 3’ elements of your choice.
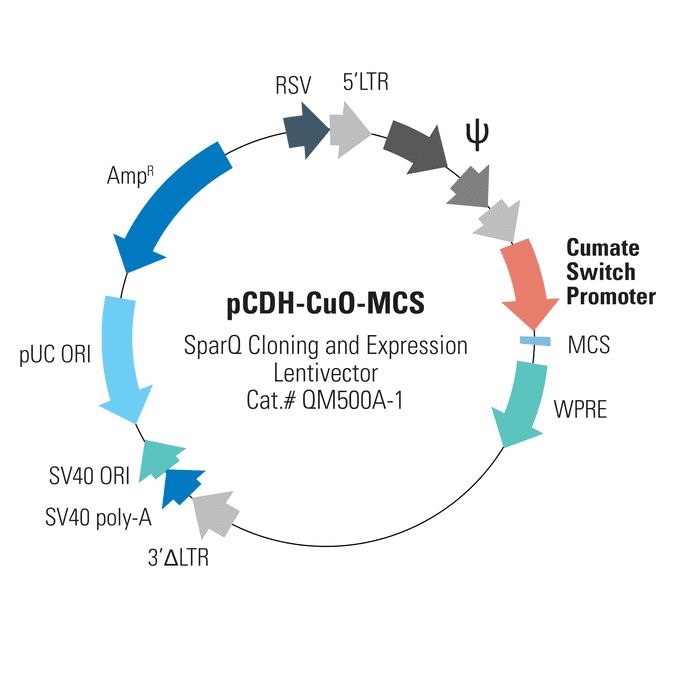 With SBI’s SparQ™ Cumate Switch System, you can get inducible gene expression in mammalian cells through the binding of cumate, a non-toxic small molecule, to CymR. CymR can be delivered to cells on its own lentivector (Cat.#s QM200PA/VA-2 and QM300PA/VA-1), or on the same lentivector as your gene-of-interest (Cat.#s QM800A-1 and QM812B-1). Expression levels of your gene-of-interest are tightly controlled and increase with increasing cumate concentration until maximum induction is reached—see as much as a 32-fold increase in gene expression. Even better, induction is reversible, so you can turn expression levels on and off. Delivering negligible background expression in the absence of cumate, the SparQ System is an excellent choice for achieving controlled levels of gene expression.
With SBI’s SparQ™ Cumate Switch System, you can get inducible gene expression in mammalian cells through the binding of cumate, a non-toxic small molecule, to CymR. CymR can be delivered to cells on its own lentivector (Cat.#s QM200PA/VA-2 and QM300PA/VA-1), or on the same lentivector as your gene-of-interest (Cat.#s QM800A-1 and QM812B-1). Expression levels of your gene-of-interest are tightly controlled and increase with increasing cumate concentration until maximum induction is reached—see as much as a 32-fold increase in gene expression. Even better, induction is reversible, so you can turn expression levels on and off. Delivering negligible background expression in the absence of cumate, the SparQ System is an excellent choice for achieving controlled levels of gene expression.
- Robust—increase expression up to 32-fold
- Adjustable—tune expression levels by titrating the amount of cumate
- Reversible—turn expression on, then off, then on again
- Versatile—choose from all-in-one formats that co-express CymR and your gene-of-interest, or two-vector systems where CymR is expressed from a different plasmid
- Powerful—suitable for in vivo applications
How It Works
How It Works
Tightly-controlled, inducible gene expression
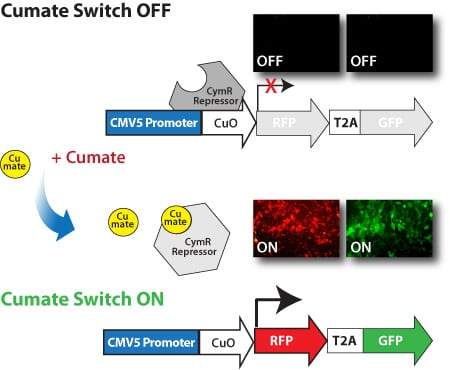
SBI’s SparQ Cumate Switch System delivers robust, titratable gene expression with low background through three components:
- Cumate, a non-toxic, small-molecule inducer
- CymR, a repressor that binds to cumate operator sequences in the absence of cumate
- SparQ Lentivector that contains an MCS to clone-in your gene-of-interest, the cumate inducible promoter with cumate operator sequences (CuO) upstream of the MCS, and one or more markers
CymR has a high binding affinity for cumate and, as more cumate is added, fewer CymR molecules bind to the CuO sequences in the promoter resulting in increased expression. Exhibiting much lower background expression than similar systems, SBI’s cumate-inducible vectors can provide up to 32-fold induction of gene expression.
Supporting Data
Supporting Data
Tight expression control with low background with the SparQ Cumate Switch System
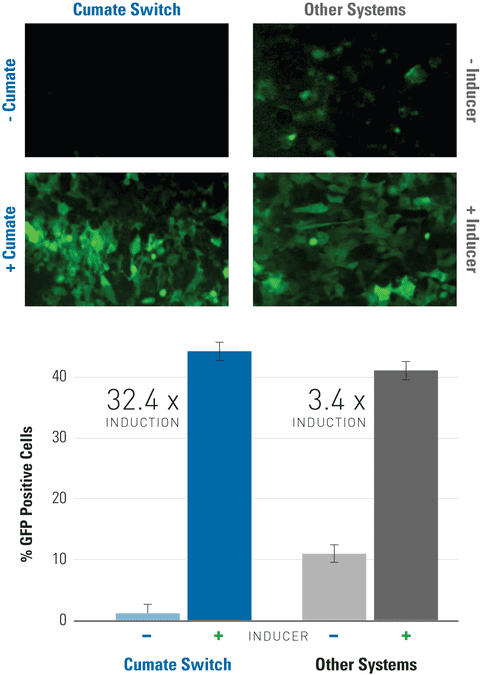
Figure 1. Get lower background and higher induction with the SparQ Cumate Switch System than other inducible systems.
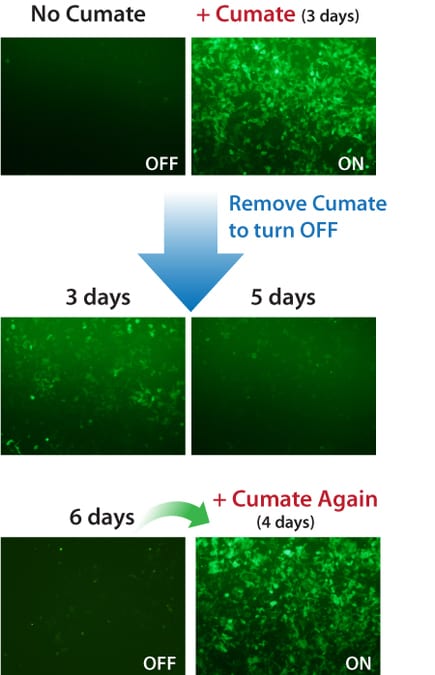
Figure 2. Gene expression with the SparQ Cumate SwitchSystem can be turned on and off, then on again.
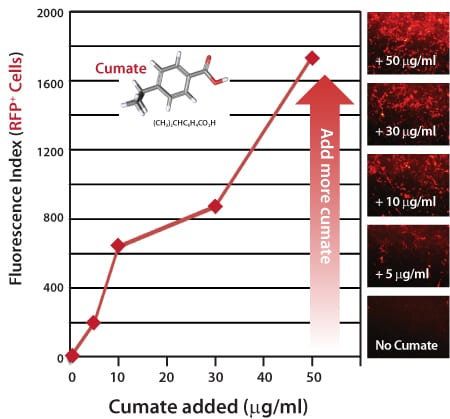
Figure 3. Gene expression with the SparQ Cumate Switch System is titratable, with increasing amounts of cumate inducing a linear increase in gene expression.
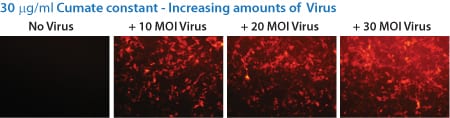
Figure 4. With the SparQ System, gene expression can also be titrated by increasing the amount of transduced SparQ lentivirus, even up to 30 MOI.
FAQs
Resources
Related Products
Citations
-
O'Hara, SP, et al. (2017) ETS Proto-oncogene 1 Transcriptionally Up-regulates the Cholangiocyte Senescence-associated Protein Cyclin-dependent Kinase Inhibitor 2A. J. Biol. Chem.. 2017; 292(12):4833-4846. PM ID: 28184004
-
Park, TS, Kim, SW & Lee, JH. (2017) Efficient transgene expression system using a cumate-inducible promoter and Cre-loxP recombination in avian cells. Asian-australas. J. Anim. Sci.. 2017; 30(6):886-892. PM ID: 27764912
-
Qi, Z, et al. (2017) An optimized, broadly applicable piggyBac transposon induction system. Nucleic Acids Res.. 2017; 45(7):e55. PM ID: 28082389
-
Shinmura, K, et al. (2017) Reduced expression of the DNA glycosylase gene MUTYH is associated with an increased number of somatic mutations via a reduction in the DNA repair capacity in prostate adenocarcinoma. Mol. Carcinog.. 2017; 56(2):781-788. PM ID: 27253753
-
Shinmura, K, et al. (2017) WDR62 overexpression is associated with a poor prognosis in patients with lung adenocarcinoma. Mol. Carcinog.. 2017; 56(8):1984-1991. PM ID: 28277612
-
Maegawa, KI, et al. (2017) The Highly Dynamic Nature of ERdj5 Is Key to Efficient Elimination of Aberrant Protein Oligomers through ER-Associated Degradation. Structure. 2017; 25(6):846-857.e4. PM ID: 28479060
-
Herrington, KA, et al. (2017) Spatial analysis of Cdc42 activity reveals a role for plasma membrane-associated Cdc42 in centrosome regulation. Mol. Biol. Cell. 2017;. PM ID: 28539409
-
Liu, L, Huang, W & Huang, JD. (2017) Synthetic circuits that process multiple light and chemical signal inputs. BMC Syst Biol. 2017; 11(1):5. PM ID: 28103878
-
Ikushima, S & Boeke, JD. (2017) New Orthogonal Transcriptional Switches Derived from Tet Repressor Homologues for Saccharomyces cerevisiae Regulated by 2,4-Diacetylphloroglucinol and Other Ligands. ACS Synth Biol. 2017; 6(3):497-506. PM ID: 28005347
-
Ando, T, et al. (2017) Ameloblastin induces tumor suppressive phenotype and enhances chemosensitivity to Dox via Src-Stat3 inactivation in osteosarcoma. Sci Rep. 2017; 7:40187. PM ID: 28054649
-
Jain, A, et al. (2017) Abl kinase regulation by BRAF/ERK and cooperation with Akt in melanoma. Oncogene. 2017;. PM ID: 28368422
-
Narumi, S, et al. (2016) SAMD9 mutations cause a novel multisystem disorder, MIRAGE syndrome, and are associated with loss of chromosome 7. Nat. Genet.. 2016; 48(7):792-7. PM ID: 27182967
-
Park, BO, et al. (2016) Selective novel inverse agonists for human GPR43 augment GLP-1 secretion. Eur. J. Pharmacol.. 2016; 771:1-9. PM ID: 26683635
-
Choi, JS, et al. (2016) cIAPs promote the proteasomal degradation of mutant SOD1 linked to familial amyotrophic lateral sclerosis. Biochem. Biophys. Res. Commun.. 2016; 480(3):422-428. PM ID: 27773815
-
Herrington, KA. (2016) New technologies for a better understanding of the Golgi: FLIM-FRET and click chemistry. Thesis. 2016;. Link: Thesis
-
Welti, J, et al. (2016) Analytical Validation and Clinical Qualification of a New Immunohistochemical Assay for Androgen Receptor Splice Variant-7 Protein Expression in Metastatic Castration-resistant Prostate Cancer. Eur. Urol.. 2016; 70(4):599-608. PM ID: 27117751
-
Barta, T, Peskova, L & Hampl, A. (2016) miRNAsong: a web-based tool for generation and testing of miRNA sponge constructs in silico. Sci Rep. 2016; 6:36625. PM ID: 27857164
-
Ushioda, R, et al. (2016) Redox-assisted regulation of Ca2+ homeostasis in the endoplasmic reticulum by disulfide reductase ERdj5. Proc. Natl. Acad. Sci. U.S.A.. 2016; 113(41):E6055-E6063. PM ID: 27694578
-
Zidek, LM, et al. (2015) Deficiency in mTORC1-controlled C/EBPβ-mRNA translation improves metabolic health in mice. EMBO Rep.. 2015; 16(8):1022-36. PM ID: 26113365
-
Peterson, MF, et al. (2015) Integrated systems for exosome investigation. Methods. 2015; 87:31-45. PM ID: 25916618
- See More
Products
| Catalog Number | Description | Size | Price | Quantity | Add to Cart | |||
|---|---|---|---|---|---|---|---|---|
| QM500A-1 | pCDH-CuO-MCS empty cloning lentivector | 10 µg | $554 |
|
||||
Overview
Overview
Get tight, titratable gene expression
With SBI’s SparQ™ Cumate Switch System, you can get inducible gene expression in mammalian cells with a range of cloning and expression lentivectors. The pCDH-CuO-MCS SparQ™ Cloning and Expression Lentivector maximizes your flexibility with only an MCS downstream of the cumate switch promoter—clone-in your gene of interest and the 3’ elements of your choice.
With SBI’s SparQ™ Cumate Switch System, you can get inducible gene expression in mammalian cells through the binding of cumate, a non-toxic small molecule, to CymR. CymR can be delivered to cells on its own lentivector (Cat.#s QM200PA/VA-2 and QM300PA/VA-1), or on the same lentivector as your gene-of-interest (Cat.#s QM800A-1 and QM812B-1). Expression levels of your gene-of-interest are tightly controlled and increase with increasing cumate concentration until maximum induction is reached—see as much as a 32-fold increase in gene expression. Even better, induction is reversible, so you can turn expression levels on and off. Delivering negligible background expression in the absence of cumate, the SparQ System is an excellent choice for achieving controlled levels of gene expression.
- Robust—increase expression up to 32-fold
- Adjustable—tune expression levels by titrating the amount of cumate
- Reversible—turn expression on, then off, then on again
- Versatile—choose from all-in-one formats that co-express CymR and your gene-of-interest, or two-vector systems where CymR is expressed from a different plasmid
- Powerful—suitable for in vivo applications
How It Works
How It Works
Tightly-controlled, inducible gene expression
SBI’s SparQ Cumate Switch System delivers robust, titratable gene expression with low background through three components:
- Cumate, a non-toxic, small-molecule inducer
- CymR, a repressor that binds to cumate operator sequences in the absence of cumate
- SparQ Lentivector that contains an MCS to clone-in your gene-of-interest, the cumate inducible promoter with cumate operator sequences (CuO) upstream of the MCS, and one or more markers
CymR has a high binding affinity for cumate and, as more cumate is added, fewer CymR molecules bind to the CuO sequences in the promoter resulting in increased expression. Exhibiting much lower background expression than similar systems, SBI’s cumate-inducible vectors can provide up to 32-fold induction of gene expression.
Supporting Data
Supporting Data
Tight expression control with low background with the SparQ Cumate Switch System
Figure 1. Get lower background and higher induction with the SparQ Cumate Switch System than other inducible systems.
Figure 2. Gene expression with the SparQ Cumate SwitchSystem can be turned on and off, then on again.
Figure 3. Gene expression with the SparQ Cumate Switch System is titratable, with increasing amounts of cumate inducing a linear increase in gene expression.
Figure 4. With the SparQ System, gene expression can also be titrated by increasing the amount of transduced SparQ lentivirus, even up to 30 MOI.
FAQs
Citations
-
O'Hara, SP, et al. (2017) ETS Proto-oncogene 1 Transcriptionally Up-regulates the Cholangiocyte Senescence-associated Protein Cyclin-dependent Kinase Inhibitor 2A. J. Biol. Chem.. 2017; 292(12):4833-4846. PM ID: 28184004
-
Park, TS, Kim, SW & Lee, JH. (2017) Efficient transgene expression system using a cumate-inducible promoter and Cre-loxP recombination in avian cells. Asian-australas. J. Anim. Sci.. 2017; 30(6):886-892. PM ID: 27764912
-
Qi, Z, et al. (2017) An optimized, broadly applicable piggyBac transposon induction system. Nucleic Acids Res.. 2017; 45(7):e55. PM ID: 28082389
-
Shinmura, K, et al. (2017) Reduced expression of the DNA glycosylase gene MUTYH is associated with an increased number of somatic mutations via a reduction in the DNA repair capacity in prostate adenocarcinoma. Mol. Carcinog.. 2017; 56(2):781-788. PM ID: 27253753
-
Shinmura, K, et al. (2017) WDR62 overexpression is associated with a poor prognosis in patients with lung adenocarcinoma. Mol. Carcinog.. 2017; 56(8):1984-1991. PM ID: 28277612
-
Maegawa, KI, et al. (2017) The Highly Dynamic Nature of ERdj5 Is Key to Efficient Elimination of Aberrant Protein Oligomers through ER-Associated Degradation. Structure. 2017; 25(6):846-857.e4. PM ID: 28479060
-
Herrington, KA, et al. (2017) Spatial analysis of Cdc42 activity reveals a role for plasma membrane-associated Cdc42 in centrosome regulation. Mol. Biol. Cell. 2017;. PM ID: 28539409
-
Liu, L, Huang, W & Huang, JD. (2017) Synthetic circuits that process multiple light and chemical signal inputs. BMC Syst Biol. 2017; 11(1):5. PM ID: 28103878
-
Ikushima, S & Boeke, JD. (2017) New Orthogonal Transcriptional Switches Derived from Tet Repressor Homologues for Saccharomyces cerevisiae Regulated by 2,4-Diacetylphloroglucinol and Other Ligands. ACS Synth Biol. 2017; 6(3):497-506. PM ID: 28005347
-
Ando, T, et al. (2017) Ameloblastin induces tumor suppressive phenotype and enhances chemosensitivity to Dox via Src-Stat3 inactivation in osteosarcoma. Sci Rep. 2017; 7:40187. PM ID: 28054649
-
Jain, A, et al. (2017) Abl kinase regulation by BRAF/ERK and cooperation with Akt in melanoma. Oncogene. 2017;. PM ID: 28368422
-
Narumi, S, et al. (2016) SAMD9 mutations cause a novel multisystem disorder, MIRAGE syndrome, and are associated with loss of chromosome 7. Nat. Genet.. 2016; 48(7):792-7. PM ID: 27182967
-
Park, BO, et al. (2016) Selective novel inverse agonists for human GPR43 augment GLP-1 secretion. Eur. J. Pharmacol.. 2016; 771:1-9. PM ID: 26683635
-
Choi, JS, et al. (2016) cIAPs promote the proteasomal degradation of mutant SOD1 linked to familial amyotrophic lateral sclerosis. Biochem. Biophys. Res. Commun.. 2016; 480(3):422-428. PM ID: 27773815
-
Herrington, KA. (2016) New technologies for a better understanding of the Golgi: FLIM-FRET and click chemistry. Thesis. 2016;. Link: Thesis
-
Welti, J, et al. (2016) Analytical Validation and Clinical Qualification of a New Immunohistochemical Assay for Androgen Receptor Splice Variant-7 Protein Expression in Metastatic Castration-resistant Prostate Cancer. Eur. Urol.. 2016; 70(4):599-608. PM ID: 27117751
-
Barta, T, Peskova, L & Hampl, A. (2016) miRNAsong: a web-based tool for generation and testing of miRNA sponge constructs in silico. Sci Rep. 2016; 6:36625. PM ID: 27857164
-
Ushioda, R, et al. (2016) Redox-assisted regulation of Ca2+ homeostasis in the endoplasmic reticulum by disulfide reductase ERdj5. Proc. Natl. Acad. Sci. U.S.A.. 2016; 113(41):E6055-E6063. PM ID: 27694578
-
Zidek, LM, et al. (2015) Deficiency in mTORC1-controlled C/EBPβ-mRNA translation improves metabolic health in mice. EMBO Rep.. 2015; 16(8):1022-36. PM ID: 26113365
-
Peterson, MF, et al. (2015) Integrated systems for exosome investigation. Methods. 2015; 87:31-45. PM ID: 25916618
- See More