Exo-FBS, heat inactivated
- Heat inactivated using a highly quality-controlled process
- Exosome-sized vesicles removed
- Very low levels of CD63-positive cow exosomes
- Undetectable levels of cow miRNAs
- Comparable growth rates as standard FBS

Products
| Catalog Number | Description | Size | Price | Quantity | Add to Cart | |||
|---|---|---|---|---|---|---|---|---|
| EXO-FBSHI-250A-1 | Exosome-depleted FBS Media Supplement Heat Inactivated - USA Certified | 250 mL | $881 |
|
||||
| EXO-FBSHI-50A-1 | Exosome-depleted FBS Media Supplement Heat Inactivated - USA Certified | 50 mL | $214 |
|
||||
Overview
Overview
Avoid inadvertently studying bovine exosomes with heat inactivated Exo-FBS
Fetal bovine serum, or FBS, is an important component of many types of cell culture media, and some researchers require heat inactivated FBS. But for researchers interested in isolating exosomes from cultured cells, standard heat inactivated FBS can introduce unwanted complications—bovine exosomes, which can cause significant background issues or interfere with functional studies. Which is why SBI developed heat inactivated Exo-FBS, our patented exosome-depleted FBS.
- Heat inactivated using a highly quality-controlled process
- Exosome-sized vesicles removed
- Very low levels of CD63-positive cow exosomes
- Undetectable levels of cow miRNAs
- Comparable growth rates as standard FBS
- Interchangeable with standard FBS (add 10% in DMEM or RPMI)
How It Works
Supporting Data
Supporting Data
High quality and great performance
Exo-FBS has greatly reduced levels of bovine exosomes (Figures 1 and 2), bovine miRNAs (Figure 3), and is even cleaner than ultracentrifuged FBS (Figure 4). Cell growth in media supplemented with Exo-FBS is similar to cell growth in media supplemented with standard FBS (Figure 5).
Exo-FBS has greatly reduced levels of bovine exosomes
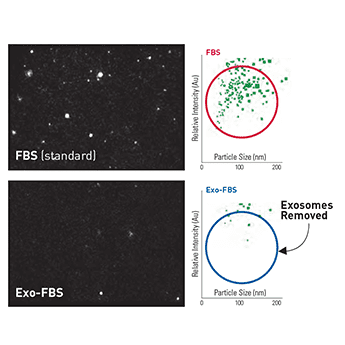
Figure 1. NanoSight particle analysis shows low levels of exosomes in Exo-FBS. While standard FBS contains exosome-sized particles (top panels), Exo-FBS shows almost no particles (bottom panels). Standard FBS and Exo-FBS samples were diluted 1:1000 and then analyzed for particle size and abundance using a NanoSight LM10 instrument.
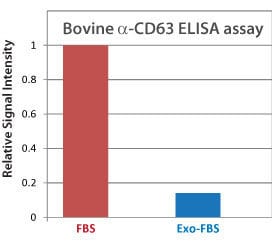
Figure 2. Bovine α-CD63 ELISA shows low levels of exosomes in Exo-FBS CD63 is an exosome-specific marker. An α-CD63 ELISA of standard FBS and Exo-FBS shows very low levels of CD63 in Exo-FBS, supporting the NanoSight particle analysis which showed low numbers of exosome-sized particles in Exo-FBS (Figure 1). Equal volumes (50 µl) of either standard FBS or Exo-FBS depleted media supplement were used and the graphed results normalized to the signal level of standard FBS.
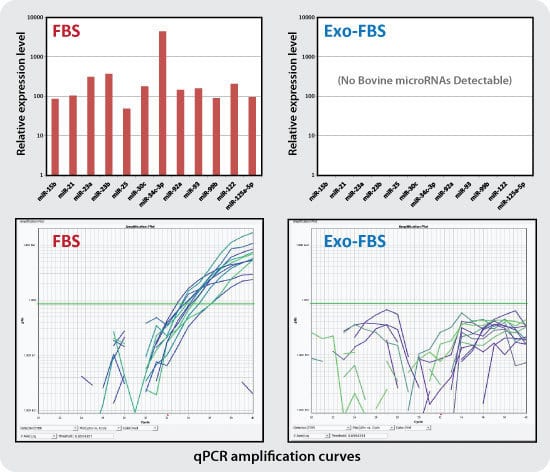
Figure 3. qPCR assays show undetectable levels of bovine exosomal miRNAs in Exo-FBS. While standard FBS contains amplifiable miRNAs (12 of the 72 individual miRNAs tested, left panels), Exo-FBS shows no amplifiable miRNAs (right panels). Standard FBS and Exo-FBS media supplements (4 ml) were treated with Trizol extraction methods to recover exosomal RNAs. RNA was converted to cDNA and 72 individual bovine microRNAs were measured by qPCR using SBI’s QuantiMir system.

Figure 4. NanoSight analysis shows Exo-FBS is cleaner than ultracentrifuged FBS. Quality Control data is generated on every batch of Exo-FBS produced at SBI by comparing NanoSight particle count analyses to the source FBS, FBS ultracentrifuged for 18 hours, and Exo-FBS. All samples were diluted 1:100 and data collected in triplicate.
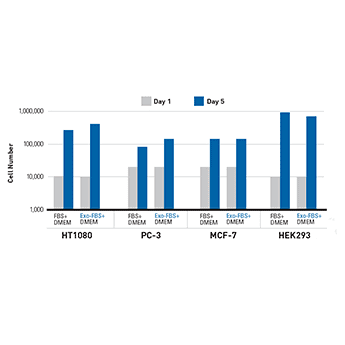
Figure 5. Cells grown in 10% Exo-FBS show comparable growth rates to 10% standard FBS. HT1080 fibrosarcoma cells, PC-3 prostate cancer cells, MCF-7 breast cancer cells, and HEK293 cells were seeded at either 10,000 or 20,000 cells and then cultured under standard conditions at 37°C with 5% CO2 for 5 days.
FAQs
Resources
Related Products
Citations
-
Liu, J, et al. (2024) Exosomes derived from impaired liver aggravate alveolar bone loss via shuttle of Fasn in type 2 diabetes mellitus. Bioactive materials. 2024; 33:85-99. PM ID: 38024229
-
Li, X, et al. (2024) The association of SPARC with hypertension and its function in endothelial-dependent relaxation. Atherosclerosis. 2024; 388:117390. Link: Atherosclerosis
-
Kang, W, et al. (2024) Lyoprotectant Constituents Suited for Lyophilization and Reconstitution of Stem-Cell-Derived Extracellular Vesicles. Biomaterials Research. 2024; 28. Link: Biomaterials Research
-
Qi, L, et al. (2024) Mesoporous bioactive glass scaffolds for the delivery of bone marrow stem cell-derived osteoinductive extracellular vesicles lncRNA promote senescent bone defect repair by targeting the miR-1843a-5p/mob3a/YAP axis. Acta Biomaterialia. 2024;. Link: Acta Biomaterialia
-
Wang, W, et al. (2024) Integrin β1-rich extracellular vesicles of kidney recruit Fn1+ macrophages to aggravate ischemia-reperfusion-induced inflammation. JCI insight. 2024; 9(2). PM ID: 38258908
-
Liu, B, et al. (2024) DNA Logical Device Combining an Entropy-Driven Catalytic Amplification Strategy for the Simultaneous Detection of Exosomal Multiplex miRNAs In Situ. Analytical chemistry. 2024; 96(4):1733-1741. PM ID: 38227423
-
Ramil, CP, et al. (2024) Extracellular vesicles released by cancer-associated fibroblast-induced myeloid-derived suppressor cells inhibit T-cell function. Oncoimmunology. 2024; 13(1):2300882. PM ID: 38192443
-
Ueda, S, et al. (2024) Consistency between Primary Uterine Corpus Malignancies and Their Corresponding Patient-Derived Xenograft Models. International Journal of Molecular Sciences. 2024; 25(3):1486. Link: International Journal of Molecular Sciences
-
Li, B, et al. (2024) Characteristics of Inflammatory and Normal Endothelial Exosomes on Endothelial Function and the Development of Hypertension. Inflammation. 2024;. PM ID: 38240985
-
Luo, P, et al. (2024) Human Umbilical Cord Mesenchymal Stem Cell-Derived Exosomes Rescue Testicular Aging. Biomedicines. 2024; 12(1). PM ID: 38255205
-
Jin, Y, et al. (2024) The homologous tumor-derived-exosomes loaded with miR-1270 selectively enhanced the suppression effect for colorectal cancer cells. Cancer medicine. 2024; 13(1). PM ID: 38197582
-
Kashiwagi, R, Udono, M & Katakura, Y. (2024) Fructobacillus fructosus OS-1010 strain stimulates intestinal cells to secrete exosomes that activate muscle cells. Cytotechnology. 2024;. Link: Cytotechnology
-
Park, DJ, et al. (2024) Defining the activity of pro-reparative extracellular vesicles in wound healing based on miRNA payloads and cell type-specific lineage mapping. Molecular therapy : the journal of the American Society of Gene Therapy. 2024;. PM ID: 38379282
-
Qian, Y, et al. (2024) M2 macrophage-derived exosomal miR-26b-5p regulates macrophage polarization and chondrocyte hypertrophy by targeting TLR3 and COL10A1 to alleviate osteoarthritis. Journal of nanobiotechnology. 2024; 22(1):72. PM ID: 38374072
-
Lv, X, et al. (2024) NSP6 inhibits the production of ACE2-containing exosomes to promote SARS-CoV-2 infectivity. mBio. 2024;:e0335823. PM ID: 38303107
-
Lee, SJ, et al. (2024) EGF-conditioned M1 macrophages Convey reduced inflammation into corneal endothelial cells through exosomes. Heliyon. 2024; 10(5):e26800. Link: Heliyon
-
Yang, LM, et al. (2024) Exosome-Transmitted miR-224-5p Promotes Colorectal Cancer Cell Proliferation via Targeting ULK2 in p53-Dependent Manner. Biomedical and environmental sciences : BES. 2024; 37(1):71-84. PM ID: 38326722
-
Wu, Y, et al. (2024) Human lens epithelial-secreted exosomes attenuate ocular angiogenesis via inhibiting microglial activation. Experimental Eye Research. 2024; 241:109837. Link: Experimental Eye Research
-
Liu, Y, et al. (2024) Extracellular Vesicles Obtained From Lung Adenocarcinoma Cells Cultured Under Intermittent Hypoxia Induce M2 Macrophage Polarization via miR-20a-5p Delivery. Technology in cancer research & treatment. 2024; 23:15330338231219415. PM ID: 38327167
-
Wu, Q, et al. (2023) Modification of adipose mesenchymal stem cells-derived small extracellular vesicles with fibrin-targeting peptide CREKA for enhanced bone repair. Bioactive Materials. 2023; 20:208-220. Link: Bioactive Materials
- See More
Products
| Catalog Number | Description | Size | Price | Quantity | Add to Cart | |||
|---|---|---|---|---|---|---|---|---|
| EXO-FBSHI-250A-1 | Exosome-depleted FBS Media Supplement Heat Inactivated - USA Certified | 250 mL | $881 |
|
||||
| EXO-FBSHI-50A-1 | Exosome-depleted FBS Media Supplement Heat Inactivated - USA Certified | 50 mL | $214 |
|
||||
Overview
Overview
Avoid inadvertently studying bovine exosomes with heat inactivated Exo-FBS
Fetal bovine serum, or FBS, is an important component of many types of cell culture media, and some researchers require heat inactivated FBS. But for researchers interested in isolating exosomes from cultured cells, standard heat inactivated FBS can introduce unwanted complications—bovine exosomes, which can cause significant background issues or interfere with functional studies. Which is why SBI developed heat inactivated Exo-FBS, our patented exosome-depleted FBS.
- Heat inactivated using a highly quality-controlled process
- Exosome-sized vesicles removed
- Very low levels of CD63-positive cow exosomes
- Undetectable levels of cow miRNAs
- Comparable growth rates as standard FBS
- Interchangeable with standard FBS (add 10% in DMEM or RPMI)
How It Works
Supporting Data
Supporting Data
High quality and great performance
Exo-FBS has greatly reduced levels of bovine exosomes (Figures 1 and 2), bovine miRNAs (Figure 3), and is even cleaner than ultracentrifuged FBS (Figure 4). Cell growth in media supplemented with Exo-FBS is similar to cell growth in media supplemented with standard FBS (Figure 5).
Exo-FBS has greatly reduced levels of bovine exosomes
Figure 1. NanoSight particle analysis shows low levels of exosomes in Exo-FBS. While standard FBS contains exosome-sized particles (top panels), Exo-FBS shows almost no particles (bottom panels). Standard FBS and Exo-FBS samples were diluted 1:1000 and then analyzed for particle size and abundance using a NanoSight LM10 instrument.
Figure 2. Bovine α-CD63 ELISA shows low levels of exosomes in Exo-FBS CD63 is an exosome-specific marker. An α-CD63 ELISA of standard FBS and Exo-FBS shows very low levels of CD63 in Exo-FBS, supporting the NanoSight particle analysis which showed low numbers of exosome-sized particles in Exo-FBS (Figure 1). Equal volumes (50 µl) of either standard FBS or Exo-FBS depleted media supplement were used and the graphed results normalized to the signal level of standard FBS.
Figure 3. qPCR assays show undetectable levels of bovine exosomal miRNAs in Exo-FBS. While standard FBS contains amplifiable miRNAs (12 of the 72 individual miRNAs tested, left panels), Exo-FBS shows no amplifiable miRNAs (right panels). Standard FBS and Exo-FBS media supplements (4 ml) were treated with Trizol extraction methods to recover exosomal RNAs. RNA was converted to cDNA and 72 individual bovine microRNAs were measured by qPCR using SBI’s QuantiMir system.
Figure 4. NanoSight analysis shows Exo-FBS is cleaner than ultracentrifuged FBS. Quality Control data is generated on every batch of Exo-FBS produced at SBI by comparing NanoSight particle count analyses to the source FBS, FBS ultracentrifuged for 18 hours, and Exo-FBS. All samples were diluted 1:100 and data collected in triplicate.
Figure 5. Cells grown in 10% Exo-FBS show comparable growth rates to 10% standard FBS. HT1080 fibrosarcoma cells, PC-3 prostate cancer cells, MCF-7 breast cancer cells, and HEK293 cells were seeded at either 10,000 or 20,000 cells and then cultured under standard conditions at 37°C with 5% CO2 for 5 days.
FAQs
Citations
-
Liu, J, et al. (2024) Exosomes derived from impaired liver aggravate alveolar bone loss via shuttle of Fasn in type 2 diabetes mellitus. Bioactive materials. 2024; 33:85-99. PM ID: 38024229
-
Li, X, et al. (2024) The association of SPARC with hypertension and its function in endothelial-dependent relaxation. Atherosclerosis. 2024; 388:117390. Link: Atherosclerosis
-
Kang, W, et al. (2024) Lyoprotectant Constituents Suited for Lyophilization and Reconstitution of Stem-Cell-Derived Extracellular Vesicles. Biomaterials Research. 2024; 28. Link: Biomaterials Research
-
Qi, L, et al. (2024) Mesoporous bioactive glass scaffolds for the delivery of bone marrow stem cell-derived osteoinductive extracellular vesicles lncRNA promote senescent bone defect repair by targeting the miR-1843a-5p/mob3a/YAP axis. Acta Biomaterialia. 2024;. Link: Acta Biomaterialia
-
Wang, W, et al. (2024) Integrin β1-rich extracellular vesicles of kidney recruit Fn1+ macrophages to aggravate ischemia-reperfusion-induced inflammation. JCI insight. 2024; 9(2). PM ID: 38258908
-
Liu, B, et al. (2024) DNA Logical Device Combining an Entropy-Driven Catalytic Amplification Strategy for the Simultaneous Detection of Exosomal Multiplex miRNAs In Situ. Analytical chemistry. 2024; 96(4):1733-1741. PM ID: 38227423
-
Ramil, CP, et al. (2024) Extracellular vesicles released by cancer-associated fibroblast-induced myeloid-derived suppressor cells inhibit T-cell function. Oncoimmunology. 2024; 13(1):2300882. PM ID: 38192443
-
Ueda, S, et al. (2024) Consistency between Primary Uterine Corpus Malignancies and Their Corresponding Patient-Derived Xenograft Models. International Journal of Molecular Sciences. 2024; 25(3):1486. Link: International Journal of Molecular Sciences
-
Li, B, et al. (2024) Characteristics of Inflammatory and Normal Endothelial Exosomes on Endothelial Function and the Development of Hypertension. Inflammation. 2024;. PM ID: 38240985
-
Luo, P, et al. (2024) Human Umbilical Cord Mesenchymal Stem Cell-Derived Exosomes Rescue Testicular Aging. Biomedicines. 2024; 12(1). PM ID: 38255205
-
Jin, Y, et al. (2024) The homologous tumor-derived-exosomes loaded with miR-1270 selectively enhanced the suppression effect for colorectal cancer cells. Cancer medicine. 2024; 13(1). PM ID: 38197582
-
Kashiwagi, R, Udono, M & Katakura, Y. (2024) Fructobacillus fructosus OS-1010 strain stimulates intestinal cells to secrete exosomes that activate muscle cells. Cytotechnology. 2024;. Link: Cytotechnology
-
Park, DJ, et al. (2024) Defining the activity of pro-reparative extracellular vesicles in wound healing based on miRNA payloads and cell type-specific lineage mapping. Molecular therapy : the journal of the American Society of Gene Therapy. 2024;. PM ID: 38379282
-
Qian, Y, et al. (2024) M2 macrophage-derived exosomal miR-26b-5p regulates macrophage polarization and chondrocyte hypertrophy by targeting TLR3 and COL10A1 to alleviate osteoarthritis. Journal of nanobiotechnology. 2024; 22(1):72. PM ID: 38374072
-
Lv, X, et al. (2024) NSP6 inhibits the production of ACE2-containing exosomes to promote SARS-CoV-2 infectivity. mBio. 2024;:e0335823. PM ID: 38303107
-
Lee, SJ, et al. (2024) EGF-conditioned M1 macrophages Convey reduced inflammation into corneal endothelial cells through exosomes. Heliyon. 2024; 10(5):e26800. Link: Heliyon
-
Yang, LM, et al. (2024) Exosome-Transmitted miR-224-5p Promotes Colorectal Cancer Cell Proliferation via Targeting ULK2 in p53-Dependent Manner. Biomedical and environmental sciences : BES. 2024; 37(1):71-84. PM ID: 38326722
-
Wu, Y, et al. (2024) Human lens epithelial-secreted exosomes attenuate ocular angiogenesis via inhibiting microglial activation. Experimental Eye Research. 2024; 241:109837. Link: Experimental Eye Research
-
Liu, Y, et al. (2024) Extracellular Vesicles Obtained From Lung Adenocarcinoma Cells Cultured Under Intermittent Hypoxia Induce M2 Macrophage Polarization via miR-20a-5p Delivery. Technology in cancer research & treatment. 2024; 23:15330338231219415. PM ID: 38327167
-
Wu, Q, et al. (2023) Modification of adipose mesenchymal stem cells-derived small extracellular vesicles with fibrin-targeting peptide CREKA for enhanced bone repair. Bioactive Materials. 2023; 20:208-220. Link: Bioactive Materials
- See More